Research: Piper


Piper (Piperaceae)
My research on Piper includes taxonomic and phylogenetic work, plant morphology, and the evolution of ant-plant associations. Piper, the genus of black pepper, has over 1000 species and is pantropical in distribution, although most species occur in Central America and the Andes of South America. Despite the presence of Piper in almost all tropical habitats, the genus remains poorly known. I am interested in Piper because the large number of species allows for the study of many questions related to speciation, diversification, and distribution, and because of the ant-plant associations found in some species. My research on Piper falls into four interconnected areas.
Systematics of Piper sect. Macrostachys
Within Piper, my research has been centered mainly on the subgroup Piper section Macrostachys (Tepe et al., 2004). The section is easily recognized, but species limits within the group remain unclear and estimates of the number of species within sect. Macrostachys range from 21 to over 100. The section has several interesting traits including evidence of recent and rapid speciation and the occurrence of symbiotic associations with ants in several species. A clear understanding of the species that comprise sect. Macrostachys and of the relationships among the species will provide an evolutionary framework for ecological and evolutionary studies.

Ant-plant associations
Five species in sect. Macrostachys are involved in varying degrees of symbiotic associations with ants, which make sect. Macrostachys an ideal group to study the evolution of ant-plant mutualisms (Tepe et al., 2004, 2007a, 2007b). The stems of most Piper species are solid, but there are some species that have (several species are found with) hollow stems that are occupied by ant mutualists. My comparative analysis of stem morphology has revealed significant structural differences between the stems of these different plant species, including several novel discoveries such as heterogeneous pith in stems before they are excavated, and lack of crystals in the portions of stems that are excavated.
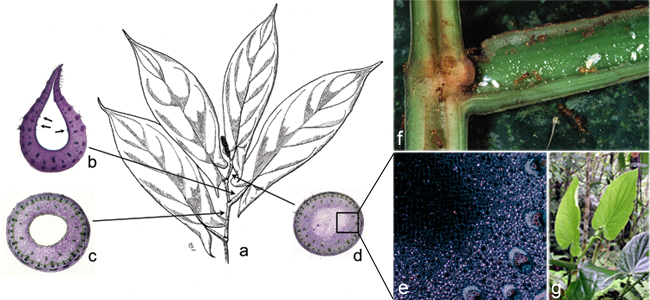
This has proven to be a fruitful line of research with several novel findings. More recently, several colleagues and I discovered and described two additional ant-Piper associations in Ecuador that differ in several important ways from those in sect. Macrostachys (Tepe et al., 2009). In addition, I have recently collected two additional hollow-stemmed species of Piper on a trip to Peru.
The systematics of Piper worldwide
Plant genera with over 1000 species, also known as “giant genera”, have generally been avoided by systematists because of their large numbers of species, taxonomic difficulty, and widespread distributions. Despite the obvious challenges, giant genera provide unique opportunities to study numerous interesting aspects of plant evolution. Using contemporary molecular techniques, tackling these giants is becoming an increasingly realistic goal for small teams of researchers. I am part of a collaborative project to provide a multi-gene phylogeny of the pantropical genus Piper. Our goal is to provide a predictive classification system for Piper to make the genus easier to use in studies of ecology, biodiversity, and conservation. The phylogeny will also be used to study floral evolution, biogeography, and the timing of divergence of major lineages (Smith et al., 2008; Jaramillo et al., 2008).

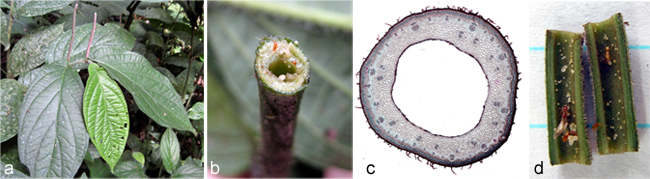
Species of New World Piper: (a) P. marginatum (sect. Pothomorphe) a common, sometimes weedy species found throughout the Neotropics; (b) P. nudilimbum, with its palmate leaf venation and globose inflorescence, is one of the most an unusual species of Piper - we don't yet know where this western Amazonian species fits into the phylogeny; (c) P. subscutatum (sect. Schilleria) is among the few Piper species with peltate leaves; and (d) the red leaf veins of P. lineolatifolium (sect. Radula) make this one of the more striking Piper species in the lowlands of Ecuador and Peru.
Patterns of evolution and diversification
A phylogenetic framework in studies of evolution is a powerful tool for the evaluation of evolutionary trends in morphology, ecology, and life history traits. My current phylogeny suggests that mutualistic associations with ants evolved independently two to four times within sect. Macrostachys (Tepe et al., 2004). Our Piper-wide phylogeny has revealed that the two ant plants from Ecuador are members of two additional sections of Piper, indicating that ant-plant associations have evolved in at least three distantly related groups within Piper. The evolution of exceptionally similar ant-associated plant traits in otherwise very different Piper species is a remarkable example of convergent evolution.
Phylogenies can also be used to test the timing of diversifications and biogeographic patterns. Piper sect. Macrostachys has a distributional pattern similar to that observed in numerous neotropical lineages, making it a model group for studying the diversification of neotropical plants. Mounting evidence indicates that, whereas the lineages themselves might be old, radiation of many Neotropical plant species are quite recent relative to the presumed age and long-term stability of the tropics (Smith et al., 2008). Preliminary analyses indicate an origin of sect. Macrostachys in the northern Andes during the Miocene-Pliocene, possibly associated with the uplift of the northern Andes and Pleistocene climatic fluctuations, with radiation of most species in the past 2.5 million years.

