RESEARCH
Thomas
J. Algeo
Professor
of Geology
University
of Cincinnati
Biocrisis at the Permian/Triassic
Boundary: Causes and Consequences
|
The
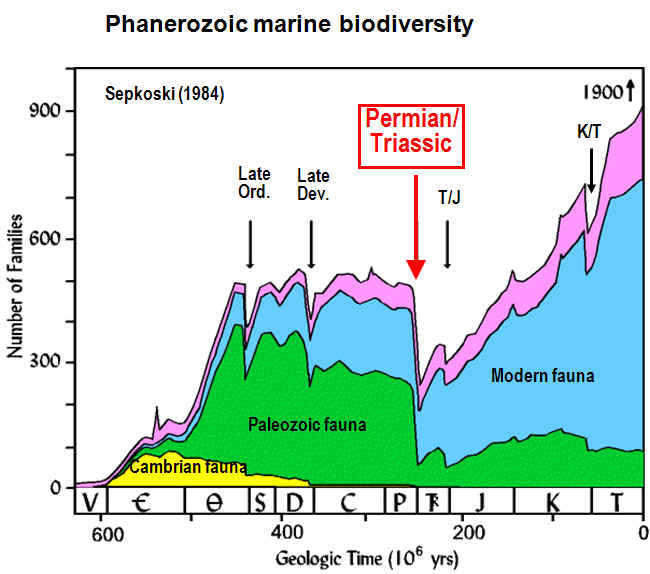
mass extinction at the Permian/Triassic boundary (PTB) was the largest
biocrisis in Earth history, eliminating ~90% of marine species and ~70% of
terrestrial species (right; Sepkoski, 2002). Eruption of the Siberian
Traps flood basalts is regarded as the most likely cause of this crisis (Renne
et al., 1995; Korte and Kozur, 2010), but the manner in which volcanic
outpourings devastated the environment and biosphere
is uncertain. I am investigating changes in environmental conditions
in marine PTB sections having a global distribution, with the goal of better
understanding the causes and consequences of this biocrisis. |
|
|
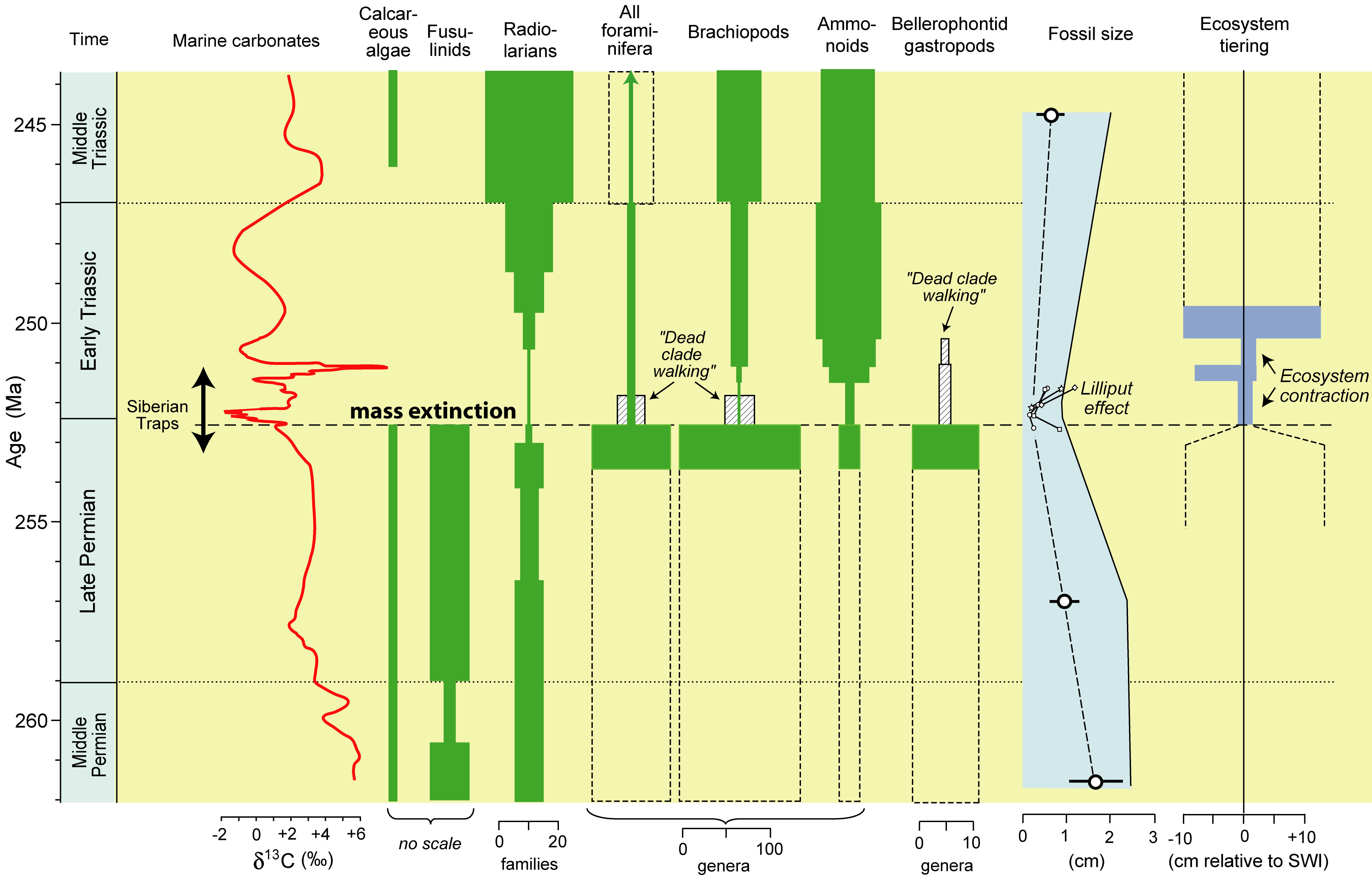
The PTB mass extinction resulted in major changes in marine
ecosystems. At Meishan (upper right), benthic marine faunas were
sharply reduced in diversity, abundance, and range of ecological tiering
from the Late Permian to the Early Triassic (Benton and Twitchett,
2003). The extinction event was broad, affecting many clades
including calcareous algae, foraminifera, radiolarians, and others (lower
right). Patterns of post-crisis recovery vary widely: some clades
quickly exceeded their pre-crisis diversity levels (e.g., ammonoids), others
were permanently reduced (e.g., brachiopods), while yet others went extinct
in the aftermath of the crisis (e.g., bellerophontid gastropods).
During the Early Triassic, some clades (e.g., bivalves and gastropods)
exhibited a pronounced reduction in average size, termed the "Lilliput
effect" (Twitchett, 2007). These ecosystem changes were
accompanied by large fluctuations in marine carbonate d13C
values (right; Payne et al., 2004), indicating major perturbations to the
global carbon cycle for several million years during the Early
Triassic. The summary figure at lower right is from Algeo et al.
(2011a). |
Late Permian
Early Triassic
|
|
|
|
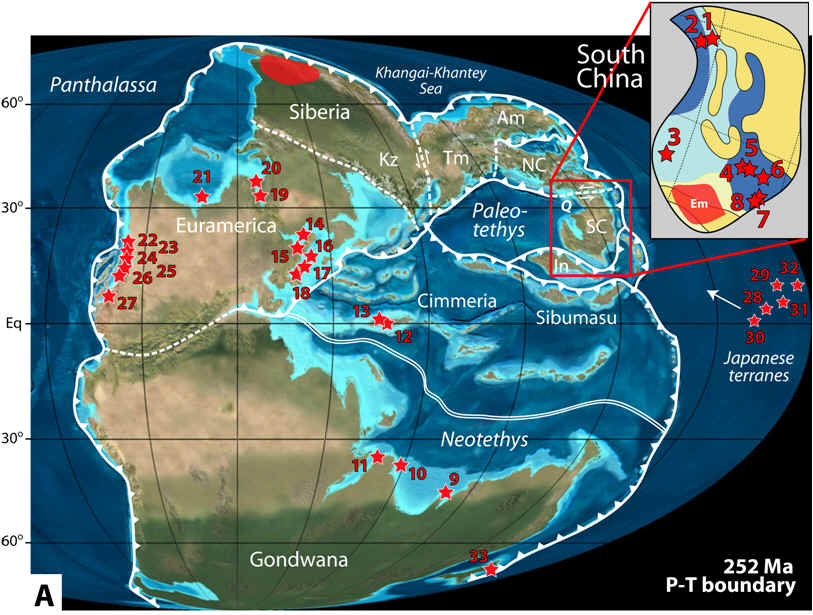
Despite decades-long study of the Permian-
Triassic boundary, there are relatively few integrated, high-resolution chemostratigraphic datasets for
marine sections that can address critical questions related to, e.g., the
extent, intensity, and timing of deep-ocean anoxia, patterns of
oxygen-minimum zone (OMZ) expansion and/or upwelling of toxic deep- ocean
waters onto shallow-marine shelves and platforms, marine-terrestrial
teleconnections,
and the relationships of these events to the delayed recovery of Early
Triassic marine biotas. My research program has generated integrated chemostratigraphic datasets for marine
sections around the world in order to address these questions (map at right with
sections numbered; adapted from Algeo et al., 2012a).
|
|
|
Earlier studies inferred that almost the
entire global ocean went anoxic during the PTB crisis (Wignall and Twitchett,
1996, 2002), and that deep-ocean anoxia commenced as early as the early Late
Permian, ~8 million years before the PTB mass extinction (Isozaki,
1997). These inferences are not in accord with the results of my
recent studies.
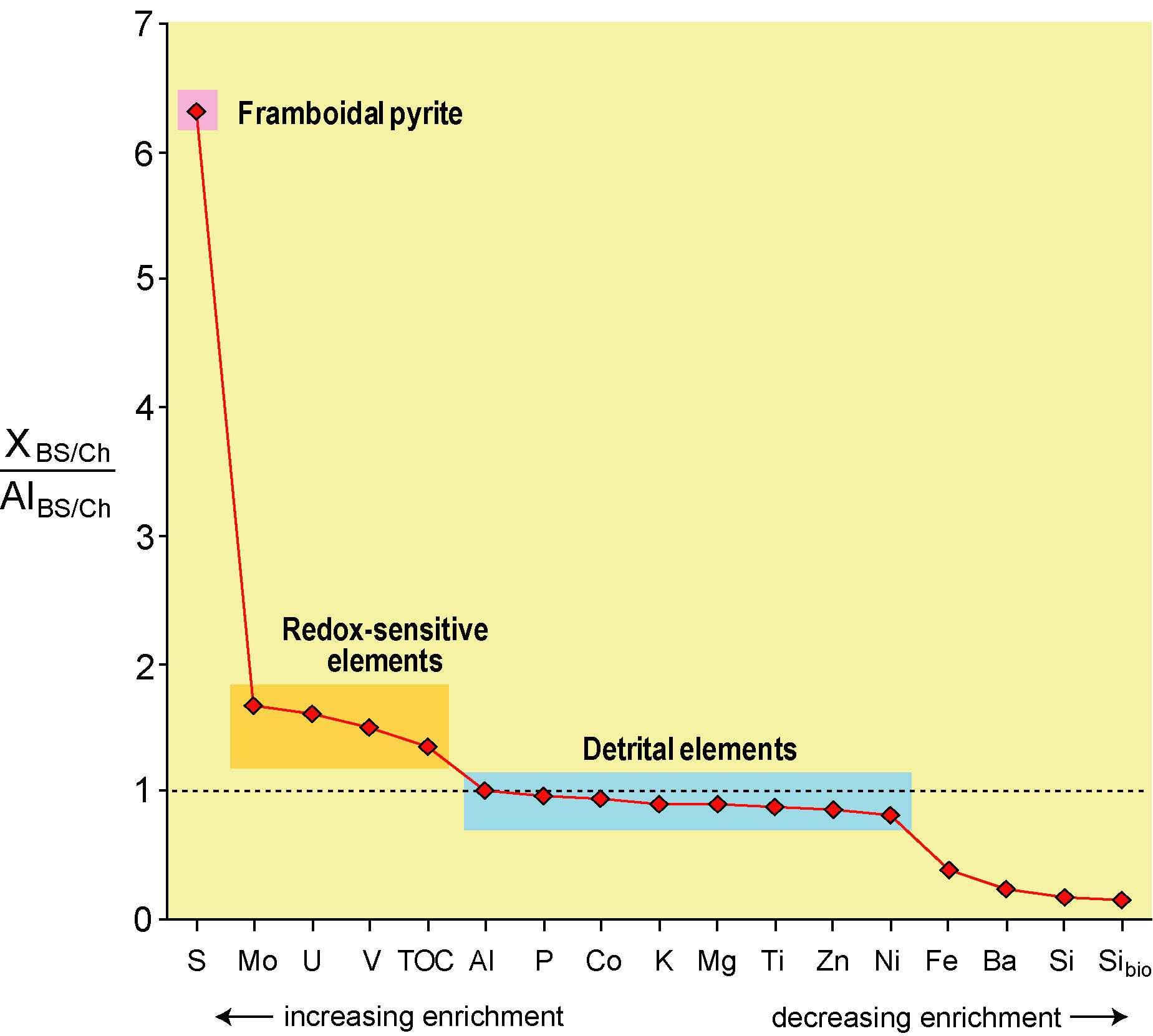
First, deep-sea sections from Japan show
evidence of only limited changes in redox conditions on the deep seafloor
(i.e., <2X increases in the concentrations of redox- sensitive elements
in the Early Triassic black shale facies relative to the Late Permian gray
chert facies; upper right, Algeo et al., 2010 and 2011b). In contrast,
there is a huge (>6X) increase in the burial flux of S related to the
appearance of pyrite framboids in the black shale facies (lower
right). I inferred that the framboids were forming high in the water
column, for example within the OMZ, rather than close to or below the
seafloor. This inference is in accord with paleoceanographic models
for the PTB (Kiehl and Shields, 2005; Winguth and Maier-Reimer, 2005;
Winguth and Winguth, 2012).
|

|
|
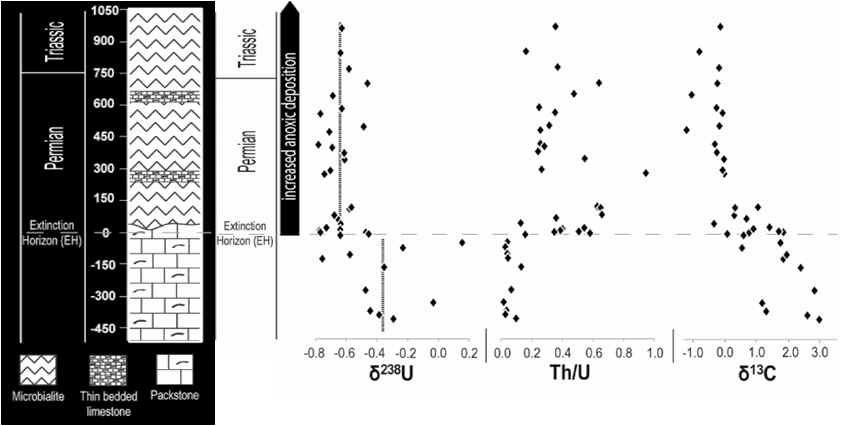
Second, the timing of changes in ocean redox
conditions during the Late Permian-Early Traissic has been extensively debated.
While small changes may have occurred in advance of the PTB crisis, U
isotope analysis of the Dawen (China) section demonstrates that a major
shift toward more reducing conditions occurred at the level of the mass
extinction horizon (Brennecka et al., 2011). This study showed that d238U
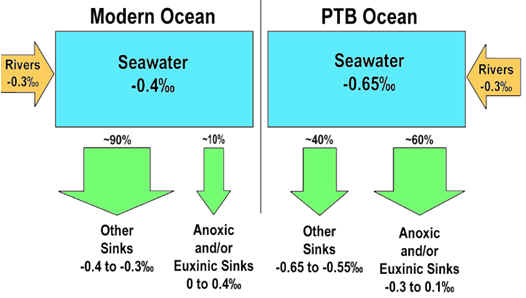
shifted rapidly from ca. -0.40‰ to ca. -0.65‰ (upper right). Mass
balance calculations indicate that this shift is consistent with expansion
of the anoxic sink for U by a factor of ~6X (from 10% to 60%; lower right).
A synchronous shift in Th/U ratios from ~0.1 to ~0.6 is also consistent with
a ~6X drawdown of U in seawater (upper right). These results
demonstrate that the most important changes in ocean redox conditions were
concurrent with the mass extinction event.
|
|
|
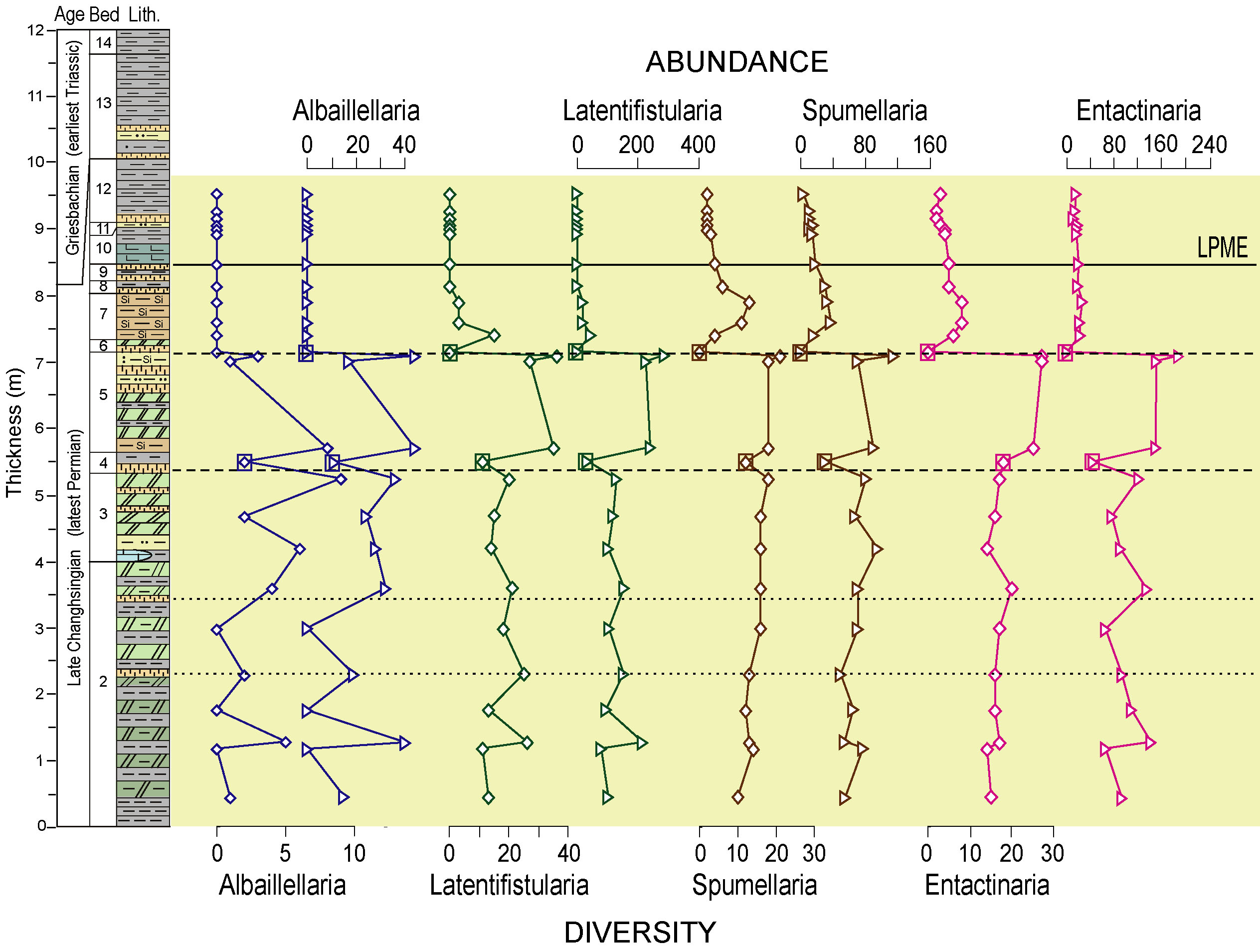
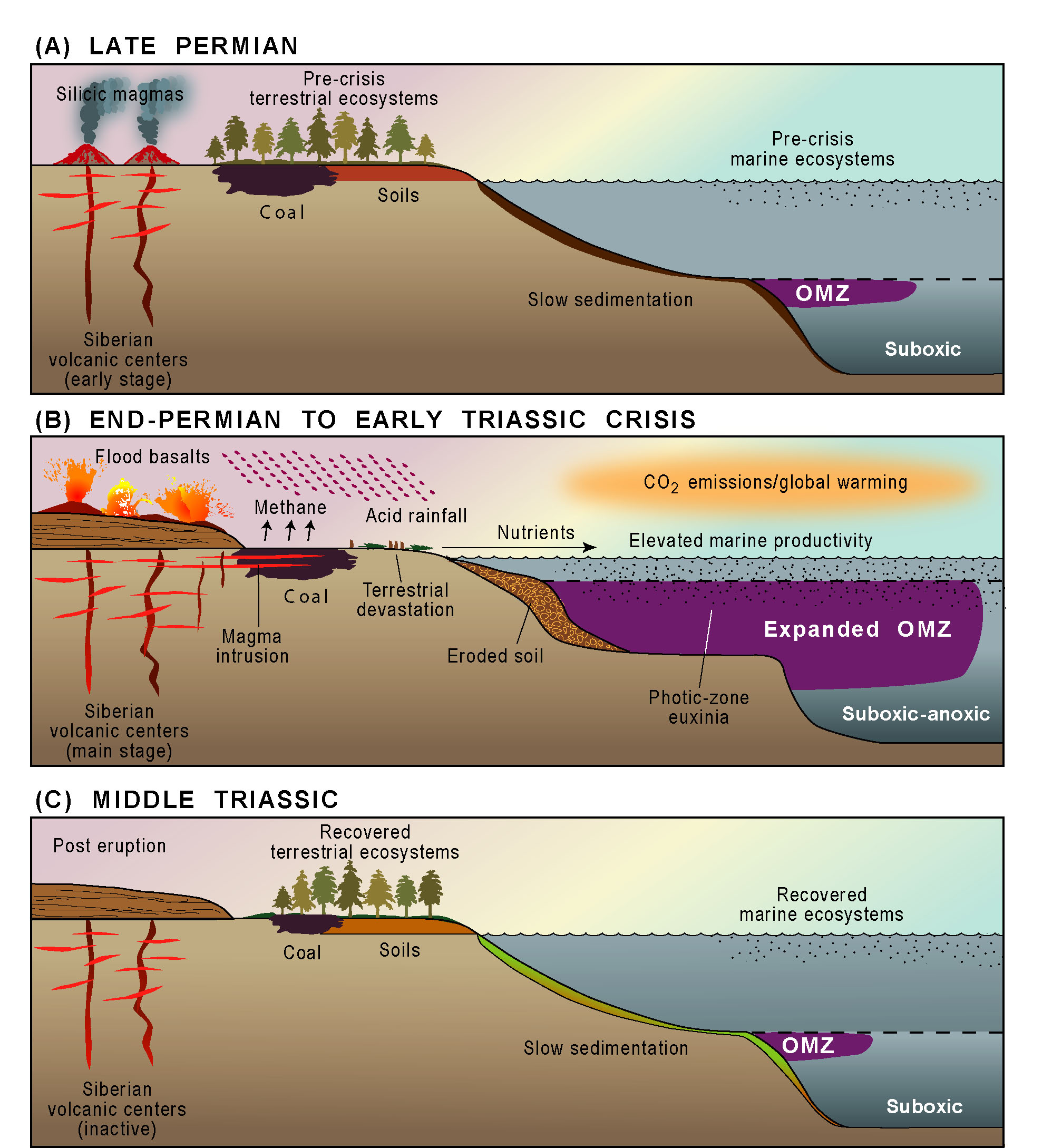
These findings suggest a major expansion of
the oceanic oxygen-minimum zone (OMZ) during the PTB crisis. Expansion
of the OMZ during the latest Permian is supported by recent analyses of
radiolarian faunas in deepwater sections of the Nanpanjiang Basin (South
China). At Dongpan, all families of radiolaria exhibit steep declines
in diversity and abundance about 2 meters below the PTB mass extinction
horizon (upper right; Shen et al., 2012a). This pattern indicates that
deepwater biota were affected by an expanding OMZ about 100kyr prior to the
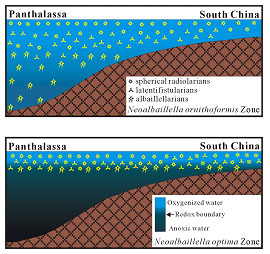
main extinction event (lower right). That radiolarian were affected by
a rising chemocline rather than by stresses imposed from the ocean surface
is demonstrated by the observation that the albaillellarian family,
representing the deepest dwelling radiolarians, went completely extinct at
this time, whereas the relatively shallower-dwelling families (Latentifistularia,
Spumelaria, and Entactinaria) declined but did not disappear (upper right).
|
|
|
 |
|
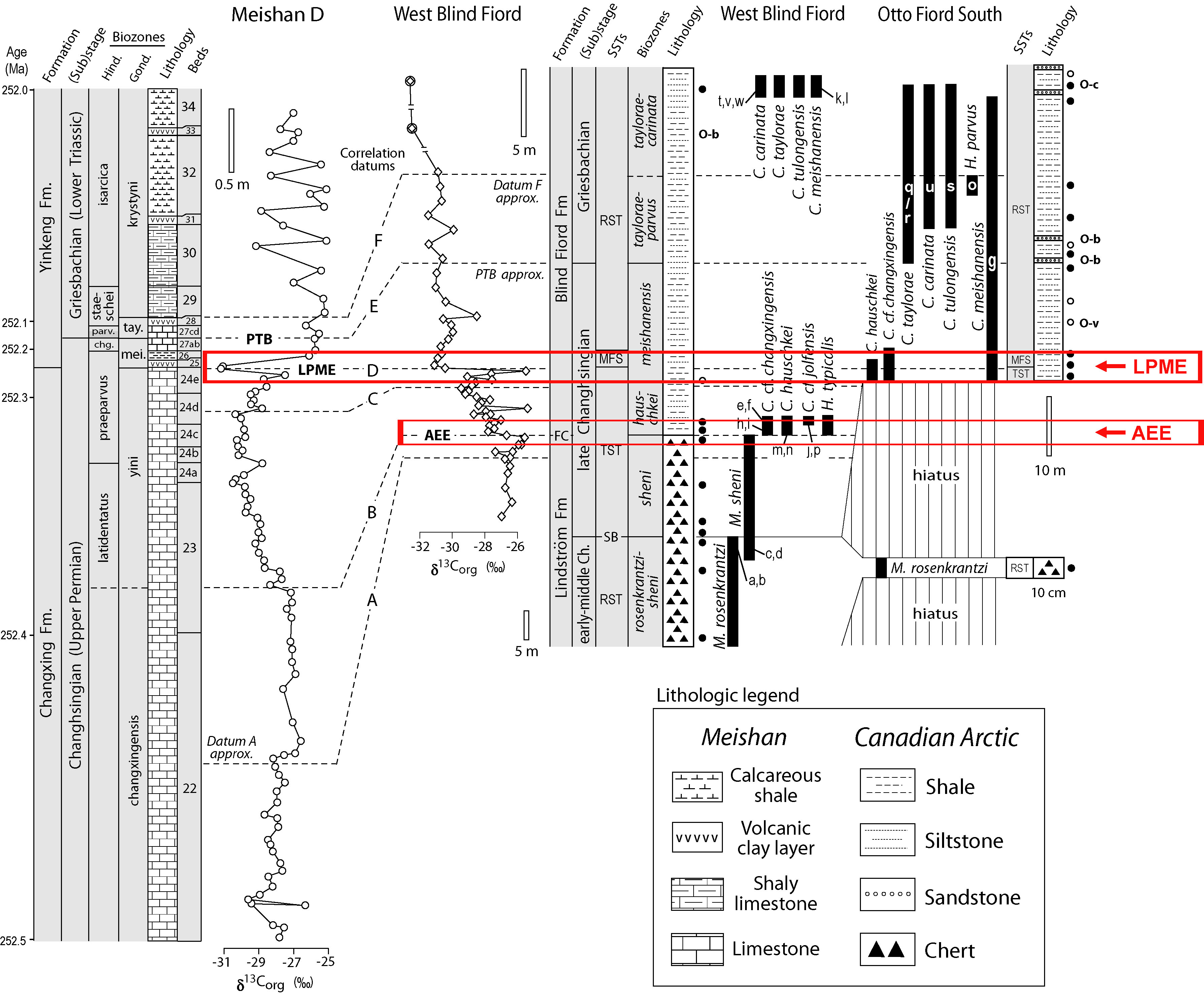
Expansion of the oceanic OMZ during the Late
Permian is implied also by biotic changes in the Sverdrup Basin of Arctic
Canada. Sections in this region such as West Blind Fiord exhibit an
abrupt extinction of siliceous sponges (the main biota of the
Late Permian Sverdrup Basin), termed the "Arctic
extinction event" (AEE) (Algeo et al., 2012b). Through detailed
conodont correlations, my
colleague, Charles Henderson of the University of Calgary, was able to show that the
AEE is older than the latest Permian mass extinction (LPME) horizon in Tethyan PTB sections (Yin et al., 2012). The implication of
this finding is that marine environmental stresses were felt earlier at high
northern paleolatitudes than in the equatorial latitudes of the Tethys
Ocean, possibly because of large-scale volcanic ash deposition or moderate
pre-crisis climatic
warming that disproportionately affected high-latitude regions (Algeo et al.,
2011a).
|
 |
|
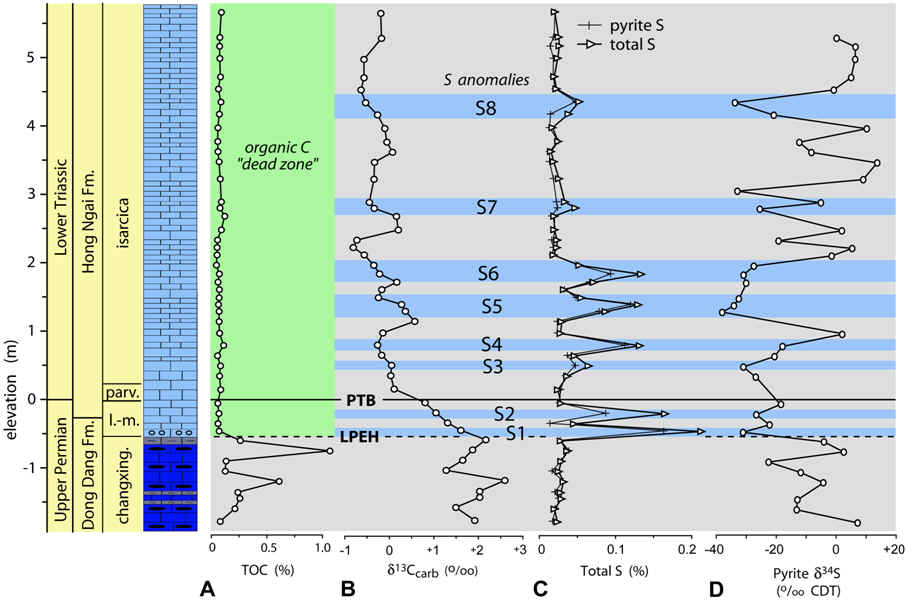
These insights regarding expansion of the
oceanic OMZ during the latest Permian help to make sense of biotic and
geochemical changes at the LPME in shallow-marine sections. PTB
sections such as that at Nhi Tao (Vietnam) accumulated on the top of a
carbonate platform in the Nanpanjiang Basin at water depths of a few meters
to tens of meters (right; Algeo et al., 2007a, 2008). Such sections
commonly show an abrupt extinction horizon that coincided with (1) a ca. 3‰
negative excursion in carbonate d13C,
(2) appearance of framboidal pyrite (S spikes) that is 34S-depleted,
and (3) near- complete loss of TOC (cf. Algeo et al., 2012a). This
pattern is consistent with episodic upward movement of sulfidic deepwaters,
possibly via the chemocline upward excursion mechanism of Kump et al.
(2005), with the first such event resulting in decimation of benthic biotas
and near-sterilization of shallow-marine habitats.
|
|
|
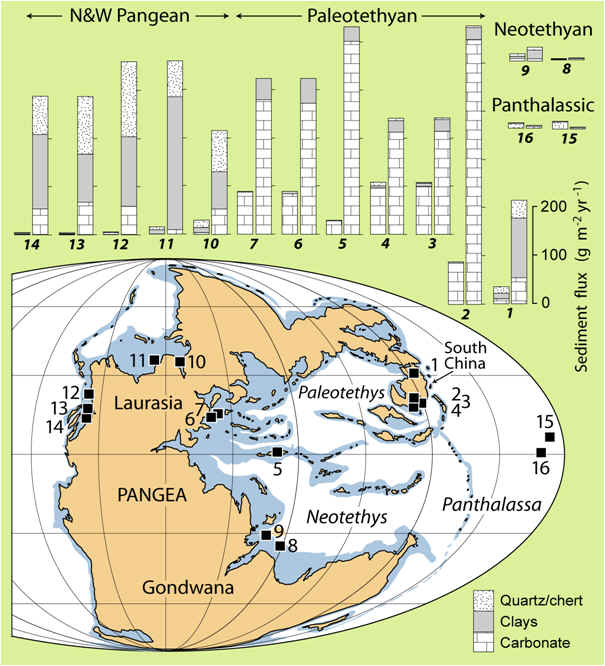
Another facet of my PTB research focuses on
terrestrial-marine "teleconnections", i.e., fluxes of material
between terrestrial and marine systems that might have played a role in
marine environmental changes during the PTB crisis. An analysis of
sediment fluxes revealed a nearly global increase in sediment accumulation
rates during the earliest Triassic (right; Algeo and Twitchett, 2010).
This increase is observed in both carbonate and siliciclastic facies in
shallow-marine areas, owing to a higher flux of dissolved and particulate
weathering products from continents, but not in deep-ocean areas that were
far from continents and below the paleo-CCD (carbonate compensation
depth). Increased continental weathering rates were probably due to a
combination of higher surface temperatures, acid rainfall, and
generally disturbed terrestrial landscapes (cf. Looy et al., 1999, 2001).
|
|
|
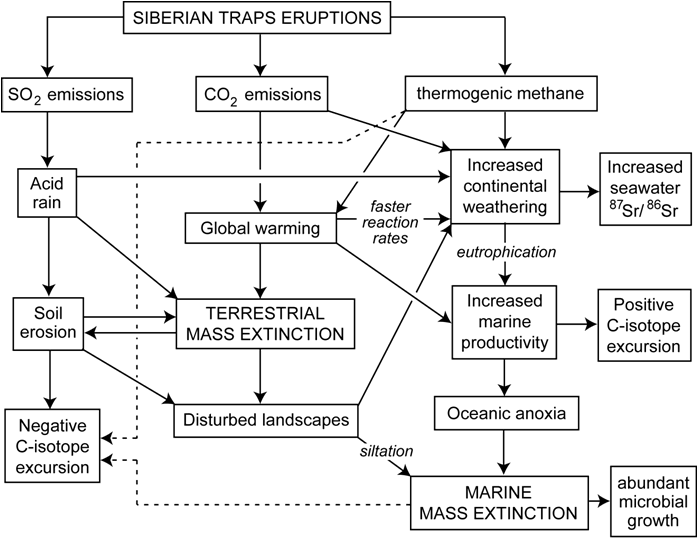
The findings above allow development of a
revised model of relationships between the Siberian Traps flood basalt
eruptions and the terrestrial and marine environmental-biotic crises (upper
right, Algeo et al., 2011a; see Wignall, 2001, for original version of
flowchart). Massive eruptions triggered strong warming through release
of volcanic CO2 and possibly also thermogenic methane following
magmatic intrusions into the West Siberian Coal Basin (lower right). A
combination of higher surface temperatures, acid rainfall, and
generally disturbed terrestrial landscapes led to an increased flux of
weathered material to shallow-marine areas. This flux included excess
nutrients that locally stimulate marine productivity, which, in combination
with warming-induced water-column stratification, resulted in a rapid
expansion of the oceanic OMZ during the latest Permian. These
conditions persisted, or recurred episodically for ~2 million years during
the Early Triassic, resulting in a delayed recovery of terrestrial and
marine ecosystems.
|
|
|
|
Work now in progress will address additional
important issues related to the Permian-Triassic boundary crisis, including:
- seawater temperature changes associated
with the crisis interval
- secular changes in seawater DIC
C-isotope gradients, and their significance for water column
stratification
- secular changes in seawater sulfate
S-isotopes, and their significance for organic burial fluxes
- secular changes in continental
weathering rates and nutrient fluxes to coastal marine systems
- the relationship of marine
environmental changes to eruption of the Siberian Traps
|
References
Algeo, T., and Twitchett, R., 2010. Anomalous Early Triassic
sediment fluxes due to due to elevated weathering rates and their biological
consequences. Geology, v. 38, p.
1023-1026. doi: 10.1130/G31203.1.
Algeo, T.J., Chen, Z.Q., Fraiser, M.L., Twitchett, R.J.,
2011a. Terrestrial-marine teleconnections in the collapse and rebuilding of Early
Triassic marine ecosystems. Palaeogeography
Palaeoclimatology Palaeoecology, v. 308, p. 1-11.
doi:10.1016/j.palaeo.2011.01.011.
Algeo, T.J.,
Ellwood, B.B., Nguyen, T.K.T., Rowe, H., and Maynard, J.B., 2007a, The
Permian-Triassic boundary at Nhi Tao, Vietnam: Evidence for recurrent influx of sulfidic watermasses to a shallow-marine
carbonate platform: Palaeogeography
Palaeoclimatology Palaeoecology, v. 252, p. 304-327,
doi:10.1016/j.palaeo.2006.11.055.
Algeo, T.J.,
Hannigan, R., Rowe, H., Brookfield, M., Baud, A., Krystyn, L., and Ellwood, B.B.,
2007b. Sequencing events across the Permian-Triassic boundary, Guryul Ravine
(Kashmir, India): Palaeogeography
Palaeoclimatology Palaeoecology, v. 252, p. 328-346,
doi:10.1016/j.palaeo.2006.11.050.
Algeo,
T., Henderson, C., Ellwood, B., Rowe, H., Elswick, E., Bates, S., Lyons, T.,
Hower, J.C., Smith, C., Maynard, J.B., Hays, L., Summons, R., Fulton, J.,
Freeman, K., 2012b. Evidence for a diachronous Late Permian marine crisis from
the Canadian Arctic region. Geological
Society of America Bulletin, v. 124 (9/10), p. 1424-1448.
doi:10.1130/B30505.1.
Algeo, T.J., Henderson, C.M., Tong, J., Feng, Q., Yin, H., Tyson, R.,
2012a.
Plankton and productivity during the Permian-Triassic boundary crisis:
An
analysis of organic carbon fluxes. Global
and Planetary Change, in press. doi:10.1016/j.gloplacha.2012.02.008.
Algeo, T.J., Hinnov, L., Moser, J., Maynard, J.B., Elswick, E., Kuwahara,
K., and Sano, H., 2010. Changes in productivity and redox conditions in the
Panthalassic Ocean during the latest Permian. Geology,
v. 38, p. 187-190. doi:10.1130/G30483.1.
Algeo, T.J., Kuwahara, K., Sano, H., Bates, S., Lyons, T., Elswick, E.,
Hinnov, L., Ellwood, B.B., Moser, J., and Maynard, J.B., 2011b. Spatial variation
in sediment fluxes, redox conditions, and productivity in the
Permian-Triassic
Panthalassic
Ocean
.
Palaeogeography Palaeoclimatology
Palaeoecology,
v. 308, p. 65-83. doi:10.1016/j.palaeo.2010.07.007.
Algeo, T.J., Shen,
Y., Zhang, T., Lyons, T.W., Bates, S.M., Rowe, H., and Nguyen, T.K.T., 2008,
Association of 34S-depleted pyrite layers with negative carbonate
δ13C excursions at the Permian/Triassic boundary: Evidence for
upwelling of sulfidic deep-ocean watermasses. Geochemistry
Geophysics Geosystems, v. 9, Q04025, 10 pp., doi:10.1029/2007GC001823.
Benton, M.J., Twitchett, R.J., 2003. How to kill
(almost) all life: the end-Permian extinction event. Trends in Ecology and
Evolution 18, 358-365.
Brennecka, G.A., Herrmann, A.D., Algeo, T.J., Anbar, A.D., 2011. Rapid
expansion of oceanic anoxia immediately before the end-Permian mass extinction. Proceedings of the National Academy of Sciences (USA), v. 108, p. 17631-17634, doi/10.1073/pnas.1106039108.
Isozaki,
Y., 1997, Permo-Triassic boundary superanoxia and stratified superocean; records
from lost deep sea: Science, v.
276, p. 235-238.
Kiehl,
J.T., and Shields,
C.A.
, 2005. Climate simulation of the latest Permian: Implications for mass
extinction. Geology v. 33, p.
757-760.
Korte,
C., Kozur, H.W., 2010. Carbon-isotope stratigraphy across the Permian-Triassic
boundary: A review. J. Asian Earth Sci. 39, 215-235.
Kump,
L.R., Pavlov, A., Arthur, M.A., 2005, Massive release of hydrogen sulfide to the
surface ocean and atmosphere during intervals of oceanic anoxia: Geology,
v. 33, p. 397-400.
Looy,
C.V., Brugman, W.A., Dilcher, D.L., Visscher, H., 1999, The delayed resurgence
of equatorial forests after the Permian-Triassic ecological crisis: Proceedings
of the National Academy of Sciences, v. 96, p. 13,857–13,862.
Looy,
C.V., Twitchett, R.J., Dilcher, D.L., van Kojnijnenberg-van Cittert, J.H.A.,
Visscher, H., 2001, Life in the end-Permian dead zone: Proceedings of the
National Academy of Sciences, v. 98, p. 7879–7883.
Luo,
G., Wang, Y., Kump, L.R., Bai, X., Algeo, T.J., Yang,
H., Xie, S., 2011b.
Nitrogen fixation prevailed simultaneously with the end-Permian marine mass
extinction and its implications. Geology, v. 39, p. 647-650. doi:10.1130/G32024.1.
Luo, G., Wang, Y., Yang, H., Algeo, T.J., Kump, L.R., Huang,
J., and Xie, S., 2011a. Stepwise and large-magnitude negative shift in δ13Ccarb
preceded the main marine mass extinction of the Permian-Triassic crisis
interval. Palaeogeography
Palaeoclimatology Palaeoecology, v. 299, p. 70-82. doi:10.1016/j.palaeo.2010.10.035.
Payne, J.L., Lehrmann, D.J., Wei, J., Orchard, M.J., Schrag, D.P., Knoll, A.H., 2004. Large
perturbations of the carbon cycle during recovery from the end-Permian
extinction. Science 305, 506-509.
Renne, P.R., Zheng, Z.C., Richards, M.A., Black, M.T., Basu,
A.R., 1995. Synchrony and causal relations between Permian–Triassic boundary
crisis and Siberian flood volcanism. Science
269, 1413-1416.
Schoepfer, S.D., Henderson, C.M., Garrison, G.H., Ward, P.D.,
Foriel, J., Selby, D., Shen, Y., Hower, J.C., Algeo, T.J., 2012. Termination of
a continent-margin upwelling system at the Permian-Triassic boundary (Opal
Creek, Alberta, Canada). Global and
Planetary Change, in press, doi:10.1016/j.gloplacha.2012.07.005.
Sepkoski, J.J., Jr., 2002. A compendium of fossil marine
animal genera. Bull. Am. Paleontol. 363, 563 pp.
Shen, J., Algeo, T.J., Hu, Q., Xu, G., Zhou, L., Feng, Q.,
2012b. Volcanism in South China during the Late Permian and its relationship to
marine ecosystem and environmental changes. Global
and Planetary Change, in press. doi:10.1016/j.gloplacha.2012.02.011.
Shen, J., Algeo, T.J., Hu, Q., Zhang, N., Zhou, L., Xia, W.,
Feng, Q., 2012c. Negative C-isotope excursions at the Permian-Triassic boundary
linked to volcanism. Geology, in press
(will appear in Nov. 2012 issue).
Shen, J., Algeo, T.J., Zhou, L., Feng, Q., Yu, J., Ellwood,
B.B., 2012a. Volcanic perturbations of the marine environment in South China
preceding the latest Permian extinction event and their biotic effects. Geobiology,
v. 10, p. 82-103. doi: 10.1111/j.1472-4669.2011.00306.x.
Twitchett, R.J., 2007. The Lilliput effect in the aftermath
of the end-Permian extinction event. Palaeogeogr. Palaeoclimatol. Palaeoecol. 252,
132-144.
Wignall,
P.B., 2001. Large igneous provinces and mass extinctions. Earth-Sci. Rev. 53,
1-33.
Wignall,
P.B., Twitchett, R.J., 1996. Oceanic anoxia and the end Permian mass extinction.
Science 272, 1155-1158.
Wignall,
P.B., Twitchett, R.J., 2002. Extent, duration, and nature of the
Permian-Triassic superanoxic event. In: Koeberl, C., and MacLeod, K.G., eds.,
Catastrophic events and mass extinctions: Impacts and beyond: Geological Society
of America Special Paper 356, p. 395-413.
Winguth,
A.M.E., Maier-Reimer, E., 2005. Causes of marine productivity and oxygen changes
associated with the Permian-Triassic boundary: A reevalution with ocean general
circulation models. Mar. Geol. 217, 283-304.
Winguth, C., Winguth, A.M.E., 2012. Simulating Permian-Triassic oceanic
anoxia distribution: Implications for species extinction and recovery. Geology
40, 127-130.
Yin, H.F., Xie, S.C., Luo, G.M., Algeo, T.J., 2012. Episodic
environmental changes at the Permian-Triassic boundary of Meishan. Episodes,
in press.
Zhao, L., Chen, Z.Q., Algeo, T.J., Chen, J.P., Chen, Y.L.,
Tong, J.
N
., Gao, S., Zhou, L., Hu, Z., and Liu, Y.S., 2012. Rare-earth element patterns
in conodont albid crowns: evidence for massive inputs of volcanic ash following
the latest Permian mass extinction? Global
and Planetary Change, in press.
Last
updated 17 Sept 2012
Back
to Research Page
Back
to Homepage